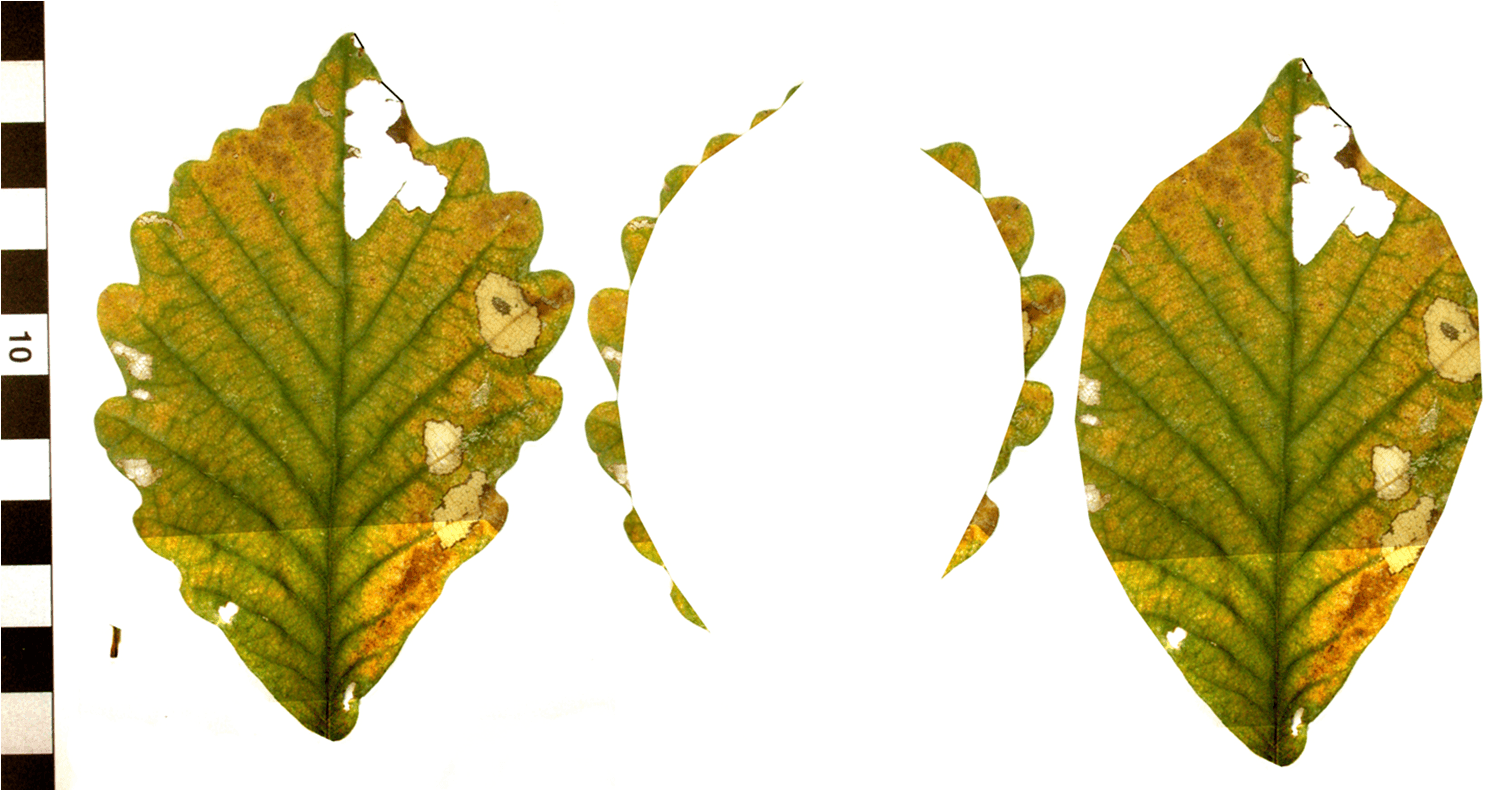|
Developing proxies for paleoclimate and paleoecological variables
If we are to develop an accurate picture of global climate during greenhouse times that can be incorporated in global climate models for past and future climate change, robust proxy data from both the terrestrial and marine records are crucial. Nonetheless, terrestrial climates during warm intervals in earth history are relatively poorly understood. Paleobotanical climate proxies that are based on the relationship between the size and shape of leaves (physiognomy) and climate are some of the best and most commonly used methodologies to reconstruct terrestrial climates from the Cretaceous through the Pleistocene.
However, these models have several shortcomings. For example, they often underestimate paleotemperatures by as much as 5 – 10 °C relative to other proxies. An underestimation of this magnitude represents the difference in reconstruction of a frost-free tropical climate and a seasonally-freezing temperate climate. Given the importance of paleobotanical climate proxies in understanding the terrestrial climate record more accurate models need to be developed and utilized in the fossil record.
|
|
Figure 1. Processed digital image of leaf. From left to right: left: Background around leaf cleaned up, margin resored, petiole removed; middle: tooth selection; right: interior of leaf with teeth removed. All digital leaf physiognomy characters are calculated using measurements derived from these images. Protocols for processing images can be found in Royer et al. (2005) and Peppe et al. (2011). (image from D. Royer) |
Digital leaf physiognomy
In an attempt to develop better paleobotanical climate models, my colleagues and I have developed multivariate models for using digital images of modern leaves (Figure 1) (Peppe et al. 2011). This method, digital leaf physiognomy, describes several leaf traits that are functionally linked to climate. We examined the relationship between physiognomy and climate at 92 globally distributed sites and found strong correlations between many leaf traits and both mean annual temperature (MAT) and mean annual precipitation (MAP) (Figure 2). Using these correlations, we developed predictive models for MAT and MAP that can be applied to fossil floras.
 |
Figure 2. Relationship between site mean of physiognomic variables and mean annual temperature for the 92 calibration sites. Standard errors of the means for each site are plotted. Linear regression fits and associated r2 and P-values are given in each panel. Figure from Peppe et al. (2011) |
Our models offer improvements in both the accuracy and precision of the climate estimates relative to traditional paleobotanical techniques. Critically, the estimates made using our new models are typically warmer and wetter, and are much closer to independent climate estimates than other leaf-climate approaches offering the potential for better understanding the terrestrial world during ancient greenhouse climates.
During this research, we noted several factors that impact the physiognomy-climate relationships such as leaf habit (deciduous vs. evergreen), local water availability, phylogenetic history, and geographic location. I am interested in exploring these factors in the future. Additionally, more calibration sites from Europe, Africa, southern South America, Oceania, and the tropics are needed to increase the phylogenetic diversity of our calibration and to explore regional differences in leaf physiognomy.
|
Paleoclimate and paleoenvironmental reconstructions in the Neogene and Quaternary in East Africa
The behavioral and morphological adaptations of mammals are a direct reflection of the habitats in which they evolve. It has been well documented that fluctuations in global climate during the Neogene and Quaternary correlate with dramatic shifts in morphologies of primate lineages. This in turn suggests that one of the driving factors in the evolution of our ancestors was paleoenvironmental change. Although this link between paleoenvironment and evolution has frequently been made, very little is known about regional climates in equatorial Africa or the paleoecology and paleoenvironments in which primate lineages evolved. Therefore, detailed reconstructions of climate and paleoenvironment are vital to better understanding the evolution of humans and their ancestors.
To explore these crucial links between paleoenvironment and the evolution of primates I am actively working on two projects in East Africa designed to reconstruct climate and paleoenvironment at several different Neogene and Quaternary hominoid fossil localities.
 |
 |
 |
| Lauren Michel, Lynsday DiPietro, and Alex Van Plantinga examining a palesol in the Hiwegi Formation on Rusinga Island, Kenya |
Tephra deposit from Pleistocene Wasiryia Beds on Rusinga Island, Kenya |
Jared Olelo and Niki Garrett collecting samples from Hiwegi Formation on Rusgina Island. |
Pleistocene paleoecology, Lake Victoria, Kenya
The Late Pleistocene history of the Lake Victoria region remains poorly understood, yet as the largest lake in Africa it is a key area for understanding the complex patterns of range expansions, contractions, and fragmentation that characterized Pleistocene plant and animal communities (including humans). I am currently collaborating on a project that is focused on understanding the paleoenvironment of equatorial East Africa during the Pleistocene and its impact on the behavioral evolution of early Homo sapiens on Rusinga Island, Lake Victoria, Kenya.
This research gives us the unique opportunity to examine both environmental change and potential human behavioral adaptations to climate changes in equatorial East Africa in a more detailed way that previously possible. Further, we will be able to relate the Pleistocene environments in the Lake Victoria region of equatorial Africa to other contemporaneous sites to test hypotheses about how environmental change affected Pleistocene hominin behavioral evolution and its impacts on modern human diversity.
Miocene paleoenvironmental reconstruction, Rusinga and Mfangano Islands, Kenya
More than 90 species of mammals, including the stem hominoid Ekembo, have been documented from Miocene deposits on Rusinga and Mfangano Islands, Kenya. These faunas form an important comparative reference for understanding the evolution of Miocene mammals in East Africa. Thus the age and paleoenvironments of the deposits are crucial to understanding Miocene mammalian paleobiology.
Our recent field studies to clarify the paleoecological and geochronological contexts of Early Miocene primate evolution have yielded new age constraints for the faunas indicating that the deposits are considerably older and represent more time than previously thought (Peppe et al., 2009; Andrews et al., 2009; Fox et al., 2009). Further, the constraints imply that the oldest hominoids from Rusinga/Mfangano may be closer in age to Proconsul species from other early Miocene localities. This suggests that taxonomic differences between the sites may be related to paleoenvironmental differences or relatively rapid (<~100 kyr) turnover in faunal composition. An exciting result of our project has been the discovery of evidence for the existence of multiple paleoenvironments in direct association of early ape fossils including open woodland habitats (Maxbauer et al. 2013) and closed-canopy forested ecosystems (Michel et al., 2014). We plan to continue working on these deposits to more fully reconstruct paleoenvironmental conditions in which these primates lived using a variety of sedimentological, paleontological, and geochemical proxies.
Miocene paleoenvironmental reconstruction and age determination, Kenya and Uganda
More recently, my research has expanded to assess the paleoenvironment and age of all of the key Early Miocene hominoid sites in Kenya and Uganda (Rusinga/Mfangano, Koru, Songhor, Karungu, Morourort, Kalodirr, Buluk, Napak, Moroto, and Bukwa). This project is part of the large collaborative REACHE project (http://www.reacheproject.com). In this interdisciplinary project, we are focused on determining the age , paleoenvironments, and mammalian fossil occurences at all of these sites across East Africa to address these major research questions:
- What types of habitats were preferred by our early ape ancestors and other primates? How did these habitats change across the East African region and through time?
- How do emerging adaptations among fossil ape species relate to environmental differences?
- How does the adaptive evolution of early Miocene apes and their relatives inform our understanding of later ape evolution and the origin of the human lineage?
|
|
Cretaceous and Paleogene plant communities and climate
Most future predictions for terrestrial habitats suggest that climate change will significantly destabilize plant and animal communities leading to extinctions, ecosystem reorganizations, and changes in biogeographic ranges. However, these predictions are based on limited paleontological data from the terrestrial world. If we are to fully understand the magnitude of terrestrial ecosystem response to changing climates, we must examine ancient environments during times of relative climate stability and compare them to times of rapid climatic change. Further, to understand how modern ecosystems will respond to extinction events, we must also understand how ancient ecosystem responded to extinctions.
The Cretaceous and Paleocene are ideal time periods to conduct this type of research for three reasons. First, they represents a time during which many modern plant and animal lineages evolved suggesting it is an excellent analog for modern and future terrestrial ecosystems. Second, they represent an interval of both climate stability and climate change. Third, they offer the opportunity to study the terrestrial ecosystem’s response to extinction.
Over the last several years, I have worked on several projects in the Northern Great Plains of North America designed to assess the evolution of plant communities through time, the terrestrial ecosystems response to the Cretaceous-Paleogene (K-Pg) extinctions, and the relationship of climate change and plant community change during the Early and Middle Paleocene.
 |
 |
| Paleocene Ginkgo leaf from Fort Union Formation, Almont, North Dakota |
“Populus” nebrascensis from Paleocene Fort Union Formation, Williston Basin, North Dakota |
Paleocene plant communities and climate
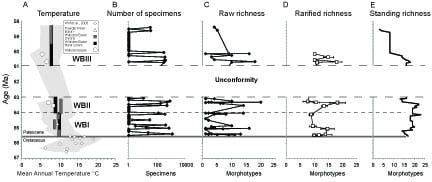
I have collected and analyzed paleomagnetic samples and fossil plants from two Paleocene sedimentary basins, the Williston and Powder River Basins, in the Northern Great Plains (Peppe, 2009; Peppe et al., 2009; Peppe, 2010). The results of my research indicate a significant drop in floral diversity from the Early to the Middle Paleocene, which may be due to a cooling climate (Peppe, 2010) (Figure 3). Based on these correlations to other basins, this drop in diversity may be regionally significant. My research also suggests that changes in the early Paleocene plant communities in North America are related both to floral evolution after the K-Pg boundary extinctions and to paleoenvironmental forcing factors.
 |
Figure 3. Estimated mean annual temperatures (A) and megafloral richness (B-D). Age estimates are based on sedimentation rates in Peppe et al. (2009) using the geomagnetic polarity time scale of Ogg and Smith (2004). Dashed lines indicate stratigraphic position of megafloral zone boundaries. White area from ~63 ~61 Ma represent depositional hiatus in the Williston Basin at the Ludlow-Tongue River formational contact. Figure from Peppe (2010). |
Although we know a great deal about the floral record in the Paleogene in the mid-latitudes in North America, much less is known about the floras in low-latitude basins, limiting our understanding of the evolution of Paleocene plant communities and their response to changing climates.
Currently I am working on a collaborative project in the San Juan Basin in New Mexico focused on documenting the Early Paleocene plant and mammalian records and correlating the records to a changing regional and global climate. This project incorporates magnetostratigraphy, sequence stratigraphy, sedimentology, paleopedology, paleobotany, vertebrate paleontology, and geochemistry. We are examining the relationship of climate change to faunal and floral change. We will also compare this climate record to the global record and to the record of faunal and floral change across North America to explore the influence of climate on early Paleocene biotic evolution.
 |
 |
 |
| Paleocene fossil leaves from San Juan Basin |
Nacimiento Formation, San Juan Basin |
Panandont jaw from the San Juan Basin |
|
|